- Филогенетическое дерево
-
Для термина «Дерево» см. другие значения.
Филогенетическое дерево (эволюционное дерево, дерево жизни) — дерево, отражающее эволюционные взаимосвязи между различными видами или другими сущностями, имеющими общего предка.
Вершины филогенетического дерева делятся на три класса: листья, узлы и (максимум один) корень. Листья — это конечные вершины, то есть те, в которые входят ровно по одному ребру; каждый лист отображает некоторый вид живых организмов (или иной объект, подверженный эволюции, например, домен белка). Каждый узел представляет эволюционное событие: разделение предкового вида на два или более, которые в дальнейшем эволюционировали независимо. Корень представляет общего предка всех рассматриваемых объектов. Ребра филогенетического дерева принято называть «ветвями».
Идея «дерева» появилась в ранних взглядах на жизнь, как на процесс развития от простых форм к сложным. Современные эволюционные биологи продолжают использовать деревья для иллюстрации эволюции, так как они наглядно показывают взаимосвязи между живыми организмами.
Содержание
Типы филогенетических деревьев
Укоренённое дерево — дерево, содержащее выделенную вершину — корень. Укоренённое дерево можно считать ориентированным графом, поскольку на нём имеется естественная ориентация — от корня к листьям. Каждый узел укоренённого дерева отвечает последнему общему предку нижележащих листьев дерева. Рисунок 1 представляет укоренённое филогенетическое дерево, окрашенное в соответствии с трёхдоменной системой живых организмов[2].
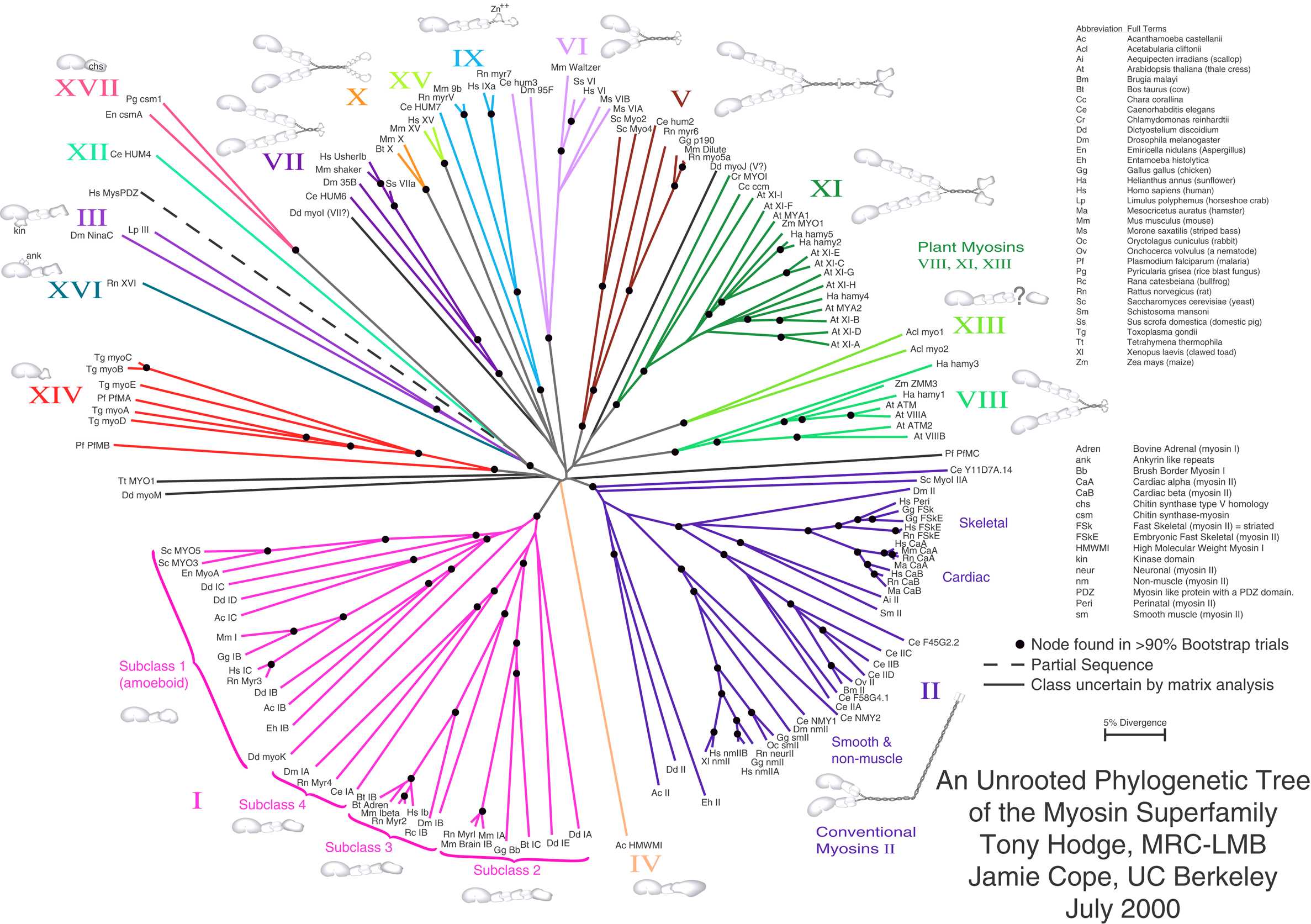
Неукоренённое дерево не содержит корня и отражает связь листьев без предполагаемого положения общего предка. Необходимость рассматривать неукоренённые деревья возникает из-за того, что часто связи между узлами восстановить легче, чем направление эволюции. Рисунок 2 представляет неукоренённое филогенетическое дерево[3]. Наиболее достоверным методом для превращения неукоренённого дерева в укоренённое (для этого надо либо объявить корнем один из узлов, либо разбить одну из ветвей на две, выходящие из корня) является использование «внешней группы» видов — достаточно близких к интересующему нас набору видов (для достоверного восстановления топологии дерева для объединенного множества видов), но в то же время заведомо являющихся отдельной группой. Иногда положение корня можно угадать, исходя из каких-либо дополнительных знаний о природе изучаемых объектов (видов, белков, etc.)
Укоренённое и неукоренённое филогенетическое дерево может быть бифуркационным или небифуркационным, а также маркированным или немаркированным. В бифуркационном дереве к каждому узлу подходят ровно три ветви (в случае укоренённого дерева — одна входящая ветвь и две исходящие). Таким образом бифуркационное дерево предполагает, что все эволюционные события состояли в происхождении от предкового объекта ровно двух потомков. К узлу небифуркационного дерева могут подходить четыре и более ветви. Маркированное дерево содержит названия листьев, тогда как немаркированное просто отражает топологию.
Дендрограмма — общий термин, обозначающий схематическое представление филогенетического дерева.
Кладограмма — филогенетическое дерево, не содержащее информации о длинах ветвей.
Филограмма (или фенограмма) — филогенетическое дерево, содержащее информацию о длинах ветвей; эти длины представляют изменение некой характеристики.
Хронограмма — филограмма, длины ветвей в которой представляют эволюционное время.
Построение филогенетических деревьев
В наше время филогенетические деревья, как правило, реконструируются по последовательностям белков или нуклеиновых кислот (ДНК или РНК). Деревья строятся по произвольному числу входных последовательностей с использованием специальных вычислительных (филогенетических) методов. Наиболее достоверной мерой соответствия данного набора (выровненных) последовательностей данной топологии дерева считается мера (критерий), основанная на принципе наибольшего правдоподобия. Нередко используется также принцип Байеса, хотя его применимость спорна[4]. Вероятностные критерии (наибольшее правдоподобие и байесовские) требуют много машинного времени для своего вычисления. Другие критерии: так называемая «максимальная экономия», а также несколько критериев, основанных на предварительном вычислении расстояний между входными последовательностями, вычисляются быстрее. Для любого критерия встаёт задача нахождения оптимального (для данной меры) дерева. Время точного решения этой задачи растёт факториально (много быстрее, чем экспоненциально!) с ростом числа входных последовательностей, поэтому для нахождения дерева, в достаточной мере удовлетворяющего входным данным, используются эвристический поиск и методы оптимизации. Кроме того, популярны такие алгоритмы, как «метод объединения соседей» (Neighbor-joining method), которые вообще не используют перебор топологий и вычисление какого-либо критерия, а сразу строят дерево, которое при хорошем качестве входных данных достаточно достоверно.
Методы построения дерева могут быть оценены по нескольким основным показателям[5]:
- эффективность (сколько времени и памяти требуются для вычислений)
- полезность (есть ли польза от полученных данных или информация бесполезна)
- воспроизводимость (будут ли повторные ответы такими же, если каждый раз даются разные данные для той же проблемной модели)
- устойчивость к ошибкам (справляется ли с неточностями в предпосылках рассматриваемой модели)
- выдача предупреждений (будет ли предупреждать нас, когда неправильно используется, то есть предпосылки неверные)
Кроме того, методы можно разделить на предполагающие и не предполагающие справедливость гипотезы «молекулярных часов» (то есть одинаковой скорости изменений для всех предковых последовательностей, см. нейтральная теория молекулярной эволюции). Первая группа методов строит укоренённые деревья, вторая — как правило, неукоренённые. Чаще всего предполагать «молекулярные часы» нет достаточных оснований, и поэтому большинство компьютерных программ филогенетического анализа по умолчанию выдают неукоренённые деревья.
Методы построения дерева могут применяться не только биологами, но и математиками. Деревья можно строить, используя Т-теорию[6].
Среди бесплатных пакетов филогенетического анализа большим набором реализованных алгоритмов выделяется PHYLIP, среди платных — PAUP. Для редактирования и визуализации готовых деревьев наиболее часто используются программы FigTree, TreeView. На русском языке доступна программа UGENE, распространяющаяся по свободной лицензии и включающая в себя построение и визуализацию деревьев.
Ограничения использования филогенетических деревьев
Хотя филогенетические деревья, построенные на основе последовательностей генов или всего генома особей различных видов, могут дать представление об эволюции, у них есть серьёзные ограничения. Филогенетические деревья не обязательно (и вероятно никогда) не дают полного и абсолютно правильного описания эволюционной истории. Данные, на которых они основываются, содержат шум. Из-за этого дерево, построенное на основе ограниченных данных (например, последовательности единственного гена или белка), требует скептического отношения к себе, потому что дерево, построенное на основе другого независимого источника данных, нередко будет отличаться от первого.
Горизонтальный перенос генов, а также гибридизация между видами затрудняют использование деревьев для описания эволюции. В случаях, когда подобные эволюционные события сыграли большую роль в эволюции изучаемой группы, предлагается использовать более сложные методы описания, например филогенетические сети (англ.).
В основном, вывод дерева филогенетического анализа — это оценка филогении особенностей (то есть дерева гена), а не филогении таксона (то есть дерева видов), из которого были отобраны эти отличительные характерные особенности, хотя в идеале оба должны быть весьма близкими.
Когда вымершие виды включены в дерево, они являются конечными точками, поскольку маловероятно, чтобы они были прямыми предками любых существующих видов. Следует скептически относиться к включению в дерево вымерших видов, информация о которых полностью или частично основана на данных цепочки ДНК, на основе того факта, что небольшая полезная «древняя ДНК» сохраняется дольше 100 000 лет, и, за исключением необычных случаев, цепочка ДНК не является достаточно длинной для использования в филогенетических анализах, даже если она взята из материала давностью до 1 млн лет.
В некоторых организмах эндосимбионты могут иметь генетическую историю, независимую от носителя.
Филогенетические сети используются, когда бифуркация деревьев не уместна, из-за этих сложностей охват эволюционной истории выбранных организмов имеет более сетчатый узор.
См. также
- Кладограмма
- Эволюционное учение
- Эволюционная биология
- Филогенетика
- Таксономия
- Генеалогическое древо
- Т-теория
Примечания
- ↑ Hodge, T. & M. J. T. V. Cope. 2000. A Myosin Family Tree. Journal of Cell Science 113: 3353-3354. See also the Myosin external link below.
- ↑ Woese, C. R. 1998. The Universal Ancestor. en:Proceedings of the National Academy of Sciences 95: 6854-6859.
- ↑ Maher, B. A. 2002. Uprooting the Tree of Life. The Scientist 16: 18 (Sep. 16, 2002); subscription only
- ↑ Felsenstein J. (2004). Inferring Phylogenies Sinauer Associates: Sunderland, MA.
- ↑ Penny, D., Hendy, M. D. & M. A. Steel. 1992. Progress with methods for constructing evolutionary trees. Trends in Ecology and Evolution 7: 73-79.
- ↑ A. Dress, K. T. Huber, and V. Moulton. 2001. Metric Spaces in Pure and Applied Mathematics. Documenta Mathematica LSU 2001: 121—139
Ссылки
Изображения в интернете
- Филогенетические деревья, основанные на 16 rDNA (англ.) (Проверено 7 ноября 2009) (недоступная ссылка)
- Страница с примером работы программы 3D View (англ.) (Проверено 7 ноября 2009)
- Филогенетическое дерево Y-хромосомы человека, 2002 г. (англ.) (Проверено 7 ноября 2009)
- «Дерево жизни» в научном журнале 2003 года (англ.) (Проверено 7 ноября 2009)
- Эволюционное дерево гоминид — предков человека (на основании палеоантропологических данных
Общее описание
- Филогенетический анализ семейства белков-гомологов (Проверено 7 ноября 2009)
- PhyloCode
- A Multiple Alignment of 139 Myosin Sequences and a Phylogenetic Tree (англ.) (Проверено 7 ноября 2009)
- Веб-проект построения «дерева жизни» (англ.) (Проверено 7 ноября 2009)
- Семейство филогенетических деревьев эволюции динозавров (англ.) (Проверено 7 ноября 2009)
 Эволюция • Доказательства эволюции
Эволюция • Доказательства эволюцииЭволюционные процессы Адаптация • Преадаптация • Экзаптация • Абаптация • Видообразование • Микроэволюция • Макроэволюция Генетика популяций Дрейф генов • Естественный отбор • Изоляция • Поток генов Происхождение жизни Возникновение жизни • Химическая эволюция • Гипотеза мира РНК Исторические концепции Дарвинизм • Ламаркизм • Пангенезис • Ортогенез • Номогенез • Сальтационизм • Катастрофизм Современные теории Синтетическая теория эволюции • Теория прерывистого равновесия • Нейтральная теория молекулярной эволюции • Эволюционная биология развития • Эпигенетическая теория эволюции Эволюция таксонов Растения • Земноводные • Рептилии • Птицы • Млекопитающие • Китообразные • Человек История эволюционного учения • Хронология эволюции • История жизни на Земле Категории:- Биологическая систематика
- Эволюционная биология
- Эволюция групп живых организмов


Wikimedia Foundation. 2010.